
Как сделать кладограмму
Добавил пользователь Дмитрий К. Обновлено: 04.10.2024
В соответствии с кладистическим подходом в группы объединяют только те виды, которые имеют общие приобретённые признаки, то есть признаки, которые отсутствовали у их дальних предков.
Кладистический подход, как полагают, является наилучшим методом для филогенетического анализа, потому что он принимает текущую эволюционную теорию и опирается на нее, то есть предполагает, что новые виды образуются при разветвлении эволюционных линий (то есть путём кладогенеза).
Кладом называют монофилетический таксон.
Клад - это группа организмов или генов, в которую входит ближайший общий предок всех её членов и все потомки этого ближайшего общего предка. Термин "клад" (англ. clade) произошёл от греческого слова κλаδоζ, означающего "ветвь", "ответвление" или "прут". Изредка в литературе этот термин встречается в женском роде - "клада".
Таксоном называют систематическую группу в классификации организмов. Узел представляет собой точку разветвления эволюционной линии. В некоторых случаях длины ветвей соответствуют степени расхождения видов (рисунок 65).

Рисунок 65 - Родство трёх млекопитающих, представленное в виде ветви филогенетического дерева
Примеры кладограмм, в которых порядок разветвлений соответствует эволюционным последовательностям видообразований у предков, представлен на рисунках 21 и 22.
Кладистический подход рассматривает возможные направления эволюции, выводит характеристики предков на всех узлах и выбирает оптимальное дерево согласно некоторой эволюционной модели. Отправной точкой кладистики служит тот факт, что члены одной группы, или одного клада, имеют общую эволюционную историю и более тесно связаны друг с другом, чем с членами любой другой группы.
Данная группа определяется по некоторым общим характерным особенностям, которые отсутствовали у далёких предков. Такими общими приобретёнными признаками могут быть любые характеристики, которые можно визуально отметить и описать. Обычно кладистический анализ проводят или по набору фенотипических признаков, или по множеству пар оснований или аминокислот в последовательности.
В кладистике приняты три основных допущения.
1. Организмы любой группы связаны между собой происхождением от общего предка.
2. Эволюционные линии периодически разветвляются.
3. С течением времени у потомков происходит изменение характеристик.
Кладистические методы имеют дело исключительно с паттернами наследования, полученными из анализа возможных деревьев таксонов. Они нацелены на выбор правильного дерева и используют детальные модели эволюционных процессов.
Наиболее популярными кладистическими методами являются:
1) метод молекулярной филогении - метод максимальной экономии (Maximum Parsimony);
2) метод наибольшего правдоподобия (Maximum Likelihood).
Эти методы ориентированы на данные о последовательностях и начинаются с множественного выравнивания. Оба метода не ориентируются на анатомические особенности организмов, такие как средний вес взрослой особи.
Метод наибольшей экономии (Maximum Parsimony или метод минимальной эволюции), предложенный Фитчем (W. Fitch), предсказывает эволюционное дерево, которое минимизирует число шагов, необходимых для воспроизведения наблюдаемого изменения в последовательностях.
Для того чтобы с наибольшей вероятностью предсказать, в каких позициях последовательностей символы (элементы последовательностей) совпадают, требуется построить множественное выравнивание этих последовательностей. В таких позициях множественного выравнивания будут расположены вертикальные столбцы с совпадающими элементами последовательностей. Для каждой выровненной позиции определяют такие филогенетические деревья, которые требуют наименьшее число эволюционных изменений, необходимых для того, чтобы произвести наблюдаемые изменения в последовательностях. Такой анализ проводят для каждой позиции в выравнивании последовательностей. Наконец, определяют те деревья, которые производят наименьшее число изменений в целом для всех позиций последовательностей.
Метод максимальной экономичности применяют для построения деревьев на основе минимального числа мутаций, необходимых для преобразования одной последовательности в другую.
Например, пусть даны виды, в которых есть гомологичные последовательности ATCG, ATGG, ТТСА, TCGA (рисунок 66).
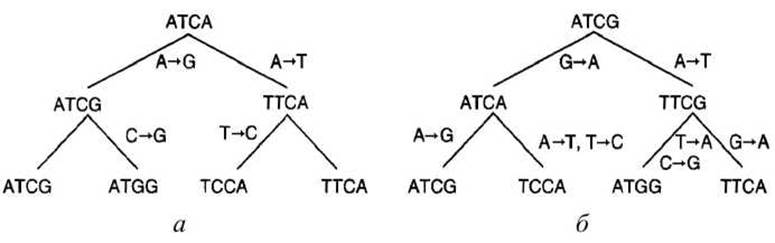
Рисунок 66 - Два генеалогических дерева мутаций
Дерево, представленное на рисунке 66(a), постулирует 4 мутации. Альтернативное дерево, представленное на рисунке 66(6), постулирует 7 мутаций. Отметим, что второе дерево постулирует, что замена G→А в четвертой позиции происходит независимо дважды.
Дерево (а) является оптимальным в соответствии с методом максимальной экономии, поскольку никакое другое дерево не даёт меньшего количества мутаций.
Во многих случаях несколько деревьев могут давать одинаковое количество мутаций, меньшее, чем другие деревья. В этих случаях метод максимальной экономии не даёт единственного ответа.
Программное обеспечение для построения филогенетических деревьев - пакет программ PHYLIP (PHYLogeny Inference Package) - состоит из 35 программ, которые не имеют графического интерфейса:
К основным средствам анализа по методу максимальной экономичности, включенным в пакет PHYLIP, относятся программы DNAPARS, DNAPENNY, DNACOMP, DNAMOVE и PROTPARS.
Метод наибольшего правдоподобия (Maximum Likelihood method), основанный на вероятностном расчёте, позволяет найти дерево, которое наилучшим образом описывает изменения, отмеченные в наборе последовательностей. Этот метод подобен методу максимальной экономичности в том, что анализ проводят для каждого столбца множественного выравнивания последовательностей. При этом учитывают все возможные деревья.
По каждому построенному дереву оценивают число вероятных изменений, или мутаций, вызвавших наблюдаемые изменения в последовательностях. Поскольку частота появления новых мутаций очень мала, постольку чем больше мутаций необходимо для приведения некоторого дерева в соответствие с имеющимися данными, тем менее правдоподобно это дерево. Для каждой топологии дерева варьируется скорость эволюции и оптимизируются параметры для того, чтобы максимизировать правдоподобие порождения наблюдаемых последовательностей.
Деревья с наименьшим количеством изменений будут наиболее правдоподобны.
Метод максимального правдоподобия позволяет построить ожидаемую модель изменений последовательности и для всех остатков взвесить вероятности их замен на любые другие остатки. Пакет PHYLIP включает в себя две программы - DNAML и DNAMLK, - предназначенные для анализа филогенетических отношений по методу максимального правдоподобия.
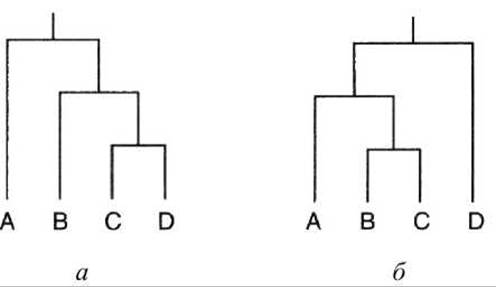
Рисунок 67 - Два филогенетических дерева мутаций
Все обсужденные выше методы не застрахованы от ошибок подобного рода, если скорость эволюции сильно варьирует на разных ветвях дерева.

Рисунок 68 - Матрицы расстояний, соответствующие филогенетическим деревьям рисунка 67
Чтобы проверить есть ли такие вариации, следует принять в рассмотрение внешнюю группу (outgroup) — виды, которые заведомо более удалены от всех видов, для которых строится дерево.
Например, если скорость эволюции у приматов постоянная, то мы ожидаем увидеть примерно одинаковые расстояния между приматами и, скажем, коровой. Если это не так, то неверно предположение о постоянстве скорости эволюции приматов.
Вычислительный анализ. Кладистические методы (максимальной экономии и наибольшего правдоподобия) более точны, чем простые методы кластеризации, такие как UPGMA, но требуют намного больше вычислительных ресурсов для решения разумных задач.
Полное количество возможных деревьев, которое должно быть просмотрено кладистическими методами, очень быстро растёт с увеличением количества видов. В результате во многих интересных случаях эти методы могут дать только приближённое решение, даже при существенных предположениях.
Поскольку вычисление филогении зачастую приближённое, то важно проверить их. Методы проверки включают:
1. Сравнение филогении полученных из разных характеристик таксонов. Являются ли они согласованными? Если деревья, полученные из разных характеристик имеют согласованные поддеревья, то они, по-видимому правильные, в то время как несогласованные поддеревья неправильные.
2. Анализ подмножеств таксонов может дать тот же ответ по отношению к подмножеству - поддерево, построенное на подмножестве должно соответствовать полному дереву.
3. Формальные статистические тесты, включающие в себя пересчёт на подмножестве исходных данных. Эти методы называются jackknife и bootstrap.
4. Если дерево имеет очень длинные ветви, то имеются серьёзные основания предполагать, что мы имеем неравномерность эволюции, а в этом случае необходимо использовать внешнюю группу.
Jackknife проводит вычисления на случайном подмножестве данных. Для построения филогении по множественному выравниванию отбираются случайные наборы позиций выравнивания и для них проводятся вычисления независимо. Если при этом восстанавливаются одинаковые поддеревья, то дерево признаётся правильным, а в противном случае - неправильным.
Bootstrap работает аналогично, но только случайно отобранные позиции могут появляться в выборке несколько раз так, чтобы размер выборки совпадал с исходной выборкой.
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Интересно то, как эволюционисты придумали способ сохранять свою гипотезу без доказательств в виде переходных форм, скрывая пробелы и создавая иллюзию реальности макроэволюции. Как они это делают? С помощью – кладограмм (схемы генеалогических связей).

Однако, кладограмма не устанавливает какой-либо генеалогической связи от предка к потомку — она всего лишь размещает организмы в соответствии со сходствами между ними. В этой диаграмме показаны в определенном порядке девять разных животных. Заметьте, что эти животные соединены друг с другом черной жирной линией, в результате чего создается впечатление, что эти группы животных эволюционировали от общего предка. На схеме из линий, напоминающей куст, показано восемь общих предков, обозначенных местами пересечения линий.
Итак, доказывает ли эта схема истинность эволюции, как заявляет Падиан? Давайте рассмотрим свидетельства. Видим ли мы свидетельства животных, показанных в конце каждой линии схемы? Да. Мы видим множество примеров лучеперых и двоякодышащих рыб, а также четвероногих животных. Кроме того, мы видим ископаемые тактаалики (род ископаемых лопастеперых рыб). Так что свидетельства существуют и расположены они на кончиках линий кладограммы или ветвей куста.
Видим ли мы свидетельства общих предков, которые обозначены пересеченными линиями кладограммы? Например, представляет ли схема свидетельства общего предка, положившего начало современным четвероногим и ископаемым тулерпетонам, как показывают пересечения их линий? Нет, этого мы не видим. А как на счет мест пересечения других линий? Нет, вообще ни одного.
Так что вся кладограмма Падиана – это не более чем расстановка живых и ископаемых животных в определенной последовательности в соответствии с различными характеристиками. По его мнению, схема доказывает, что эволюция – это факт. Но он верил в это еще до того, как составил свою схему.
Методологию кладограммы можно применить и к столовым приборам. Мы можем разложить ножи, вилки, ложки, лопатки и т.д. по определенной схеме, и это все будет выглядеть очень эффектно. Но это абсолютно не доказывает, что у ложек и ножей общий предок. Можно взять, например, такие транспортные средства, как велосипеды, мотоциклы, автомобили, грузовики, полуприцепы, и применить к ним эту же схему. Кладограмма также прекрасно сработала бы, но это не означает, что велосипеды и мотоциклы произошли от общего предка.
Кто-нибудь знает как создать кладограмму на WP? (пример в фото)
Вопрос конечно про модуль или решение. Готового не нашел, кроме редакторов диаграмм и инфографик
Но там делаются картинки, а нужно чтобы было html - адаптивность, картинки и текст ссылками
(версткой и ручками много что можно сделать, это понимаю)
Штука нужна для автора у которого нет опыта HTML, а преподавать надо - свой сайт тоже держать
Буду благодарен всем наводкам и подсказкам
- Кладограмма (англ. cladogram) — одно из основных понятий в современной биологической систематике — древовидный граф, отражающий отношения сестринского родства между таксонами.
Связанные понятия
Базальная группа (в филогенетике) — группа организмов, линия которой отделилась от корня раньше других групп. Одна клада является базальной группой по отношению к другой кладе, если количество плезиоморфных (исходных) черт у первой клады больше, чем у второй.
Игуанодонты (лат. Iguanodontia) — группа орнитоподных динозавров, живших на всех континентах планеты с нижней юры по окончание мела (164—66,0 млн лет назад).
Теропо́ды, или хищные динозавры (лат. Theropoda, буквально — звероногие, от др.-греч. θήρ — зверь и πούς — нога) — один из подотрядов ящеротазовых динозавров. Все тероподы перемещались на двух ногах, большинство — хищники, реже всеядные или растительноядные (теризинозавры, орнитомимиды). Большинство учёных с недавних пор стали относить к тероподам и современных птиц, таким образом птицы из потомков динозавров стали относиться к одному из подотрядов. Кисть обычно трёхпалая, стопа — четырёхпалая с.
Архоза́вры (лат. Archosauria, от др.-греч. ἄρχω — правлю, начинаю и σαῦρος — ящерица) — группа диапсидных пресмыкающихся, представленная в настоящее время крокодилами, а из вымерших организмов также: динозаврами, птерозаврами и другими. В кладистике в число архозавров включают птиц.
Целуроза́вры (лат. Coelurosauria) — самая многочисленная группа динозавров-тероподов, живших с середины юрского по конец мелового периода (167—66,0 млн лет назад).
Орнитоподы (лат. Ornithopoda, от др.-греч. ὄρνις, род. п. ὄρνιθος — птица и др.-греч. πούς — нога) — инфраотряд птицетазовых травоядных динозавров. Жили от верхнего юрского периода до конца мелового периода.
Парафили́я (от др.-греч. παρά — рядом и φυλή — фила) — понятие таксономии, применимое к таким классификационным системам, в которых основным критерием группировки классифицируемых объектов в таксоны оказывается степень их родства, т.е. близости к общему предку. При этом парафилетическими группами называют группы, включающие лишь часть потомков общего предка. Более формальное определение гласит: парафилетическая группа получается из монофилетической путём изъятия из состава последней одной или нескольких.
Птицетазовые (устар. птицетазовые динозавры, лат. Ornithischia) — один из двух отрядов динозавров. Несмотря на своё название, не они, а ящеротазовые динозавры по одной из версий были предками птиц.
Синапоморфия — в биологической систематике: сходство нескольких сравниваемых групп по производному состоянию признака. Родственное понятие аутапоморфия (автапоморфия) означает уникальное производное состояние признака, присущее некой группе в целом.
Валидное название (от фр. valide — важный, действующий) — в зоологической номенклатуре название, которое используется для таксона согласно правилам Международного кодекса зоологической номенклатуры (МКЗН). В ботанической номенклатуре — правильное, действительное название таксона, осуществлённое с соблюдением правил Международного кодекса ботанической номенклатуры.
Ящерота́зовые (устар. ящеротазовые динозавры, лат. Saurischia) — один из двух главных отрядов динозавров. Отличаются тем, что лобковые кости первично наклонены вперёд (позднее у некоторых представителей — сегнозавров, дромеозавров и других — лобковые кости отклоняются назад).
Теризиноза́вры (лат. Therizinosauria) — инфраотряд динозавров-тероподов из группы Maniraptoriformes, включающий в себя одно надсемейство теризинозавроидов (Therizinosauroidea) и несколько родов. Окаменелости теризинозавров были найдены в отложениях мелового периода на территории Монголии, Китая и западной части США. Различные особенности передних конечностей, черепа и таза позволяют объединить их с тероподами манирапторами — близкими родственниками птиц, несмотря на внешнее несходство.
Зауропо́ды или завропо́ды (лат. Sauropoda, буквально — ящероногие динозавры) — большая группа четвероногих растительноядных динозавров из отряда ящеротазовых обитавших с юрского по меловой периоды (около 200—66 миллионов лет назад) на всех материках; включала около 130 видов, до 13 семейств, около 70 родов. Все зауроподы вымерли. Наиболее известными представителями зауропод являются диплодоки, брахиозавры, апатозавры.
Гипсилофодонтиды (лат. Hypsilophodontidae) — небольшие динозавры из группы орнитоподов, известны из Азии, Австралии, Европы, Северной Америки и Южной Америки, из пород со средней юры до позднего мелового периода. Рассматриваются как быстрые, двуногие, растительноядные, около 1—2 метров длиной. Группа традиционно включает почти всех двуногих птицетазовых динозавров, кроме игуанодонтов.
Цера́топсы или рогатые динозавры (лат. Ceratopsia, от др.-греч. κέρας — рог и ὤψ — лицо) — группа (инфраотряд) травоядных птицетазовых динозавров верхнего юрского и мелового периодов, обладавших характерным крючковатым клювом. Окаменелые останки обнаружены в Северной Америке, Азии и Европе. Из наиболее известных представителей — цератопс и трицератопс.
Фосси́лии (лат. fossilis — ископаемый, окаменелость в палеонтологии) — ископаемые остатки организмов или следы их жизнедеятельности, относящиеся к прежним геологическим эпохам.
Цератозавры (лат. Ceratosauria) — инфраотряд тероподовых динозавров. В настоящее время нет общепринятого перечня видов цератозавров, хотя анатомически они менее разнообразны, чем тетануры.
Верхний мел (поздний мел) — вторая и последняя эпоха мелового периода. Начался 100,5 миллионов лет назад и закончился 66,0 миллионов лет назад. Продолжался, таким образом, 34,5 млн лет.
Анкилозавры, или панцирные динозавры (лат. Ankylosauria, от названия рода Ankylosaurus) — инфраотряд наземных динозавров отряда птицетазовых, характерной особенностью которых являются костные образования на туловище.
Крокодиломорфы (лат. Crocodylomorpha) — надотряд пресмыкающихся, включающий современных и ископаемых крокодилов и близкие к ним группы (крокодилоподобные архозавры, Notosuchia и другие).
Филогене́тика, или филогенети́ческая система́тика, — область биологической систематики, которая занимается выявлением и прояснением эволюционных взаимоотношений среди разных видов жизни на Земле, как современных, так и вымерших. Эволюционная теория утверждает, что сходство тех или иных особей или видов часто указывает на общее происхождение или общего предка. Потому взаимоотношения, установленные филогенетической систематикой, часто описывают эволюционную историю видов и их филогенез, исторические.
Овирапторозавры (лат. Oviraptorosauria) — инфраотряд ящеротазовых динозавров из группы целурозавров, населявших территории нынешних Северной Америки, Южной Америки, Европы и Азии. Впервые была предложена Р. Барсболдом в 1976 году.
Птицехво́стые, также орниту́ры, орниту́рные (лат. Ornithurae), — клада птиц, которой могут придавать ранг подкласса, куда включают общего предка ихтиорнисов, гесперорнисов и веерохвостых птиц, а также всех их потомков.
Птерода́ктили (лат. Pterodactyloidea, от греч. πτερόν — крыло и δάκτυλος — палец) — подотряд вымерших рептилий из отряда летающих ящеров (птерозавров), живших в юрском и меловом периодах (167,7—66,0 млн лет назад).
Синапсѝды (лат. Synapsida), также тероморфы или зверообразные (лат. Theromorpha, от др.-греч. θήρ, θηρίον — зверь + μορφή — форма) — таксономическая группа в составе клады амниот, возникшая около 318 млн лет назад. Одна из групп синапсид — цинодонты (Cynodontia) — была предком млекопитающих.
Четвероно́гие, или назе́мные позвоно́чные (лат. Tetrapoda) — надкласс позвоночных из группы челюстноротых (Gnathostomata). Характерный признак четвероногих — наличие четырёх конечностей, обычно служащих для передвижения по суше. В некоторых группах конечности существенно видоизменились для полёта (птицы, рукокрылые, птерозавры), плавания (ластоногие, китообразные, сирены, мозазавры, ихтиозавры, плезиозавры) или редуцировались (безногие земноводные, змеи, некоторые ящерицы).
Тероцефалы (лат. Therocephalia) — подотряд синапсид из отряда териодонтов, хотя некоторые систематики повышают ранг до отряда. Очень примитивные териодонты, возможно, родственные горгонопсам с одной стороны и цинодонтам — с другой. Вместе с цинодонтами образуют группу эутериодонтов (Eutheriodontia).
Архозавроморфы (лат. Archosauromorpha) — инфракласс пресмыкающихся из подкласса диапсид. Впервые появились в поздней перми и распространились в триасе. В этот инфракласс включают непосредственно архозавров (включая Crocodylia, динозавров, птиц) и некоторые вымершие отряды. В 1991 году была опубликована работа, в которой архозавроморфы рассматривались как клада, включающая наиболее раннего общего предка групп Prolacerta, архозавров, Trilophosaurus, Hyperodapedon и всех их потомков. В 1998 Дэвид Дилкс.
Карноза́вры (лат. Carnosauria) — основные хищники мезозойской эры, крупные хищники, любители свежего мяса, пожиравшие своих более слабых сородичей.
Диапсиды(лат. Diapsida) — большая группа (подкласс) завропсидов или рептилий, сформировавшаяся около 300 миллионов лет назад (в верхнем каменноугольном периоде) и существующая до наших дней (крокодилы, клювоголовые, чешуйчатые и черепахи). Следует учитывать, что в современных системах классификации птицы также относятся к диапсидам. Ныне в мире насчитывается около 14600 видов диапсидов, что делает их самой разнообразной группой высших позвоночных.
Мегалоза́вр (лат. Megalosaurus, от др.-греч. μεγάλος — огромный и σαῦρος — ящер) — род хищных двуногих динозавров из семейства Megalosauridae, живших с окончания триасового периода по конец мелового периода (208,5—66,0 млн лет назад). Ископаемые остатки найдены в Европе (Бельгия, Великобритания, Германия, Испания, Польша, Португалия и Франция), Азии (Китай) и Африке (Танзания). Мегалозавр является первым описанным динозавром.
Археопте́рикс (лат. Archaeopteryx, от греч. ἀρχαῖος — древний, πτέρυξ — крыло) — вымершее позвоночное позднего юрского периода, величиной с ворону, занимающее по морфологии промежуточное положение между пресмыкающимися и птицами, обитавшее 150—147 млн лет назад. Все находки археоптерикса относятся к окрестностям Зольнхофена на юге Германии. Долгое время (до появления других находок) использовался для реконструкции облика предполагаемого общего предка птиц.
Нижний мел — первый отдел меловой системы. Объединяет породы, отложившиеся в течение раннемеловой эпохи, которая началась около 145,0 миллионов лет назад и закончилась 100,5 миллионов лет назад (охватывая, таким образом, 44,5 млн лет).
Дицинодо́нты (лат. Dicynodontia) — клада растительноядных синапсид из подотряда аномодонтов, которой придают ранг от семейства до отряда. Жили в пермском — начале юрского периода (298,9—190,8 млн лет назад) по всему миру.
Рептилиоморфы (лат. Reptiliomorpha) — в кладистике — клада четвероногих, имеющих характеристики как рептилий, так и амфибий.
Стегоцефа́лы, или панцирноголовые (лат. Stegocephalia) — вымершая группа земноводных животных, одних из первых позвоночных, вышедших в конце девонского периода на сушу. Предками стегоцефалов были костные рыбы, обладающие дополнительными органами дыхания в виде лёгочных мешков. Наиболее близки к стегоцефалам кистепёрые рыбы, скелет которых имеет много сходных черт со скелетом стегоцефалов. Стегоцефалы вымерли в начале мезозоя.
Читайте также: